Будова листової платівки. Показані палісадна (зверху, щільно упаковані клітини) та губчаста (знизу, рихло розташовані клітини) частини мезофілу, розташовані між верхнім та нижнім епідермальними шарами
Як правило, лист складається з наступних тканин:
- Епідерміс- шар клітин, які захищають від шкідливого впливусередовища та зайвого випаровування води. Часто поверх епідермісу лист покритий захисним шаром воскоподібного походження (кутикулою).
- Мезофіл, або паренхіма- внутрішня хлорофілоносна тканина, що виконує основну функцію – фотосинтез.
- Мережа жилок, утворених провідними пучками, що складаються з судин і ситоподібних трубок, для переміщення води, розчинених солей, цукрів та механічних елементів.
- Устьиця- Спеціальні комплекси клітин, розташовані в основному на нижній поверхні листя; через них відбувається випаровування води та газообмін.
Епідерміс
Рослини в помірних та північних широтах, а також у сезонно-сухих кліматичних зонах можуть бути листопадними, тобто їхнє листя з приходом несприятливого сезону опадає або відмирає. Цей механізм має назву скиданняабо опадання. На місці опалого листа на гілочці утворюється рубець - листовий слід. В осінній період листя може забарвитися в жовтий, помаранчевий або червоний колір, оскільки зі зменшенням сонячного світла рослина зменшує вироблення зеленого хлорофілу, і лист набуває забарвлення допоміжних пігментів, таких як каротиноїди та антоціани.
Жилки
Жилки листа є судинною тканиною та розташовані в губчастому шарі мезофілу. По малюнку розгалуження жилки, зазвичай, повторюють структуру розгалуження рослини. Жилки складаються з ксилеми - тканини, що служить для проведення води та розчинених у ній мінеральних речовин, та флоеми - тканини, що служить для проведення органічних речовин, що синтезуються листям. Зазвичай ксилема лежить поверх флоеми. Разом вони утворюють основну тканину, яку називають серцевиною листа.
Морфологія листа

Хвоя їли канадської ( Picea glauca)
Основні типи листя
- Листоподібний відросток у певних видів рослин, таких як папороті.
- Листя хвойних дерев, мають голкоподібну чи шиловидную форму (хвоя).
- Листя покритонасінних (квіткових) рослин: стандартна форма включає прилисток, черешок і листову пластинку.
- Плауноподібні ( Lycopodiophyta) мають мікрофілове листя.
- Обгорткове листя (тип, що зустрічається у більшості трав)
Розташування на стеблі
У міру зростання стебла листя розташовується на ньому в певному порядку, що обумовлює оптимальний доступ до світла. Листя з'являється на стеблі по спіралі, як за годинниковою стрілкою, так і проти неї, під певним кутом розбіжності. У куті розбіжності помічена точна послідовність Фібоначчі: 1/2, 2/3, 3/5, 5/8, 8/13, 13/21, 21/34, 34/55, 55/89. Така послідовність обмежена повним оборотом в 360 °, 360 ° x 34/89 = 137,52 або 137 ° 30 "- кут, в математиці відомий під назвою золотий кут. У послідовності номер дає кількість оборотів до того моменту, поки лист не повернеться в Нижченаведений приклад показує кути, при яких листя розташоване на стеблі:
- Чергові листи розташовані під кутом 180 ° (або 1/2)
- 120° (або 1/3): три аркуші в обороті
- 144° (або 2/5): п'ять листків за два обороти
- 135° (або 3/8): вісім листків за три оберти
Зазвичай листорозташування описується за допомогою наступних термінів:
- Чергове(Послідовне) - листя розташовуються по одному (у чергу) на кожен вузол.
- Супротивне- Листя розташовуються по два на кожному вузлі і зазвичай перехресно-попарно, тобто кожен наступний вузол на стеблі розгорнуті відносно попереднього на кут 90 °; або двома рядами, а то й розгорнутий, але є кілька вузлів.
- Мутчасте- Листя розташовуються по три і більше на кожному вузлі стебла. На відміну від супротивного листя, у мутовчатих кожен наступний завиток може перебувати під кутом 90° від попереднього, а може і не перебувати, обертаючись на половину кута між листям у завитку. Слід проте врахувати, що супротивне листя може здатися мутовчастим на кінці стебла.
- Розеточне- Листя, розташовані в розетці (пучок листя, розташованих по колу з одного загального центру).
Сторони аркуша
У будь-якого листа в морфології рослин є дві сторони: абаксіальна та адаксіальна.
Абаксіальна сторона(Від лат. ab- «Від» та лат. axis- «вісь») - сторона бокового органу втечі (аркуша або спорофілу) рослини, звернена при закладці від конуса наростання (вершини) втечі. Інші назви - спинний бік, дорзальний бік.
Протилежна їй сторона називається адаксіальний(Від лат. ad- «К» і лат. axis- "Вісь"). Інші назви - черевна сторона, вентральна сторона.
У переважній більшості випадків абаксіальна сторона - це поверхня листа або спорофілу, звернена до основи втечі, проте зрідка сторона, що закладається абаксіально, розгортається в процесі розвитку на 90 або 180 і розташовується паралельно поздовжньої осі втечі або звертається до його вершини. Це характерно, наприклад, для хвої деяких видів ялини.
Терміни «абаксіальний» і «адаксіальний» зручні тим, що дозволяють описувати структури рослин, використовуючи саму рослину як систему відліку і не вдаючись до двозначних позначень типу «верхня» або «нижня» сторона. Так, для пагонів, спрямованих вертикально вгору, абаксіальна сторона бічних органів буде, як правило, нижньою, а адаксіальна - верхньою, проте якщо орієнтація втечі відхиляється від вертикальної, то терміни верхня і нижня сторона можуть ввести в оману.
Поділ листових платівок
Після того, як листові пластинки розділені, можуть бути описані дві основні форми листя.
- Простий листскладається з єдиної листової пластинки та одного черешка. Хоча він може складатися з кількох лопат, проміжки між цими лопатами не досягають основної жилки листа. Простий лист завжди опадає цілком.
- Складний листскладається з кількох листочків, розташованих на загальному черешку (який називається рахіс). Листочки, крім своєї листової пластинки, можуть мати і свій черешок (який називається черешок, або вторинний черешок). У складному аркуші кожна платівка опадає окремо. Так як кожен листочок складного листка можна розглядати як окремий листок, при ідентифікації рослини дуже важливо визначити місцезнаходження черешка. Складне листя є характерним для деяких вищих рослинтаких як бобові .
- У пальчастих(або лапчастих) листя всі листові пластинки розходяться по радіусу від закінчення корінця подібно до пальців руки. Головний черешок листа відсутній. Прикладами такого листя може бути коноплі ( Cannabis) та кінський каштан ( Aesculus).
- У перистихлистя листові пластинки розташовані вздовж основного черешка. У свою чергу, перисте листя може бути непарноперистими, з верхівковою листовою пластинкою (приклад - ясен, Fraxinus); і парноперистими, без верхівкової пластинки (приклад - червоне дерево, Swietenia).
- У двоперистихлистя листя розділені двічі: пластинки розташовані вздовж вторинних черешків, які у свою чергу прикріплені до головного черешка (приклад - альбіція, Albizzia).
- У трилистихлистя є тільки три пластинки (приклад - конюшина, Trifolium; бобовник, Laburnum)
- Перстонадрізнілистя нагадує перисті, але пластинки у них не повністю розділені (приклад - деякі горобини, Sorbus).
Характеристики черешків
Черешковілистя має черешок - стеблинку, до якого вони кріпляться. У щитовиднихлистя черешок прикріплений усередині від краю пластинки. Сидячіі обвиваючілистя черешка не мають. Сидяче листя кріпиться прямо до стебла; у обвиває листя листова пластинка повністю або частково обволікає стебло, так що складається враження, що втеча росте прямо з листа (приклад - Клейтонія пронзённолистная, Claytonia perfoliata). У деяких видів акації, наприклад, у виду Acacia koa, черешки збільшені та розширені та виконують функцію листової пластинки - такі черешки називають філлодії. Наприкінці філодію нормальний лист може існувати, а може й ні.
Характеристики прилистка
Прилистник, Присутній на листі багатьох дводольних рослин, є придатком на кожній стороні основи черешка і нагадує маленький листок. Прилистки можуть опадати зі зростанням листа, залишаючи по собі рубець; а можуть і не опадати, залишаючись разом із листом (наприклад, так відбувається у рожевих та бобових).
Прилистки можуть бути:
- вільні
- зрощені - злиті з основою черешка
- розтрубоподібні - у вигляді розтруба (приклад - ревінь, Rheum)
- оперізують основу черешка
- міжчерешкові, між черешками двох супротивних листків
- міжчерешкові, між черешком і протилежним стеблом
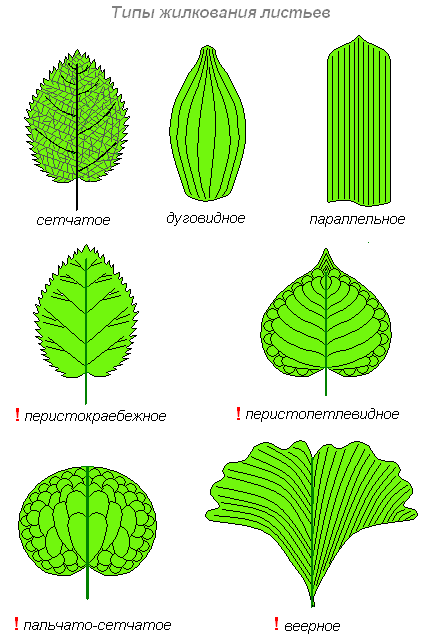
Жилкування
Існує два підкласи жилкування: крайове (основні жилки доходять до кінців листя) та дугоподібне (основні жилки проходять майже до кінців країв листа, але повертають, не доходячи до нього).
Типи жилкування:
- Сітчасте - локальні жилки розходяться від основних подібно до пір'їнка і розгалужуються на інші маленькі жилки, таким чином створюючи складну систему. Такий тип жилкування типовий дводольних рослин. У свою чергу сітчасте жилкування поділяється на:
- Перисто-нервове жилкування - лист має зазвичай одну основну жилку і безліч дрібніших, що відгалужуються від основної і йдуть паралельно один до одного. Приклад - яблуня ( Malus).
- Радіальне - лист має три основні жилки, що походять від його основи. Приклад - червонокоренник, або цеанотус ( Ceanothus).
- Пальчасте – кілька основних жилок радіально розходяться недалеко від основи черешка. Приклад - клен ( Acer).
- Паралельне - жилки йдуть паралельно вздовж усього аркуша, від його основи до кінчика. Типово для однодольних рослин, таких як злаки ( Poaceae).
- Дихотомічне – домінуючі жилки відсутні, жилки поділяються на дві. Зустрічається у гінкго ( Ginkgo) та деяких папоротей.
Термінологія листа

Термінологія опису листа

Листя з різними формами. За годинниковою стрілкою з правого кута: потрійний лопатевий, овальний з дрібнопилчастим краєм, щитовидний з пальчастим жилкуванням, загострений непарноперистий (в центрі), перисторозсічений, лопатевий, овальний з цілокраїм краєм
Форма листа
- Голчаста: тонка та гостра
- Загострена: клиноподібна з довгою вершиною
- Двоякопериста: кожен лист перистий
- Серцеподібна: у вигляді серця лист кріпиться до стебла в районі ямочки.
- Клиноподібна: лист трикутний, лист кріпиться до стебла на вершині.
- Дельтоподібна: лист трикутний, кріпиться до стебла в основі трикутника
- Пальчаста: листок поділений на пальцеподібні лопаті
- Овальна: лист овальний, з коротким кінцем
- Серпоподібна: у вигляді серпа
- Віялоподібна: напівкругла, або у вигляді віяла
- Стрілоподібна: лист у вигляді наконечника стріли, з лопатями, що розширюються, в основі
- Ланцетна: лист довгий, широкий посередині
- Лінійна: лист довгий та дуже вузький
- Лопатева: з кількома лопатями
- Назад серцеподібна: лист у вигляді серця, кріпиться до стебла на виступаючому кінці
- Назад ланцетоподібна: верхня частина ширша, ніж нижня
- Назад яйцеподібна: у вигляді сльози, лист кріпиться до стебла на виступаючому кінці
- Округла: круглої форми
- Овальна: лист овальний, яйцеподібний, із загостреним кінцем у основі
- Долоніста: розділена на багато лопатей
- Щитовидна: лист закруглений, стебло кріпиться знизу
- Периста: два ряди листочків
- Непарнопериста: лист перистий з верхівковим листочком
- Парноперистий: лист перистий без верхівкового листочка
- Перисторозсічена: лист розсічений, але не до середини
- Ниркоподібна: лист у формі нирки
- Ромбоподібна: лист у формі ромба
- Лопатоподібна: лист у вигляді лопати
- Списоподібна: гостра, з колючками
- Шилоподібна: у вигляді шила
- Трійчаста: листок поділений на три листочки
- Тричі периста: кожен листочок у свою чергу ділиться на три
- Однолопатеве: з одним листочком
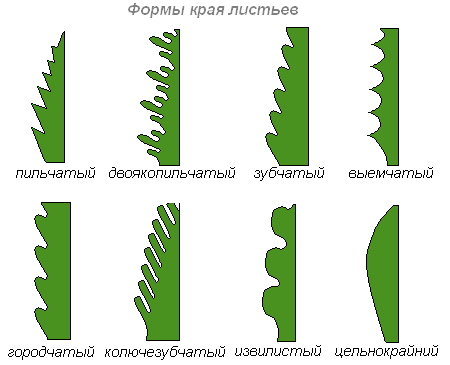
Край листа
Край листя часто є характеристикою роду рослини та допомагає визначити вид:
- Цільнокрайний - з гладким краєм, без зубців
- Реснитчастий - з бахромою по краях
- Зубчастий - із зубчиками, як у каштана. Крок зубчика може бути великий і маленький
- Округлозубчастий – з хвилеподібними зубцями, як у бука.
- Дрібнозубчастий – з дрібними зубчиками
- Лопатевий - порізаний, з вирізами, що не досягають середини, як у багатьох дубів
- Пильчастий - з несиметричними зубчиками, спрямованими вперед у бік верхівки листа, як у
Листя буває прості та складні. Складнимлистом називається такий, черешок якого має кілька листових платівок. Вони прикріплюються до головного черешка своїми власними черешками, нерідко самостійно, поодинці, відпадають і називаються листочками.Прикладами складного листа є лист конопель, люпину, конюшини, кінського каштана, волоського горіха. Простийлист має одну платівку. Від простого до складного листа існують різноманітні перехідні форми, що розрізняються за характером та ступенем порізаності пластинки.
Просте листя по обрису платівки буває овальними, яйцеподібними, зворотнояйцеподібними, ниркоподібними, довгастими, ланцетними, мечоподібними, лінійнимита ін.
Якщо краї листової пластинки не мають жодних виїмок, лист називають цілокрайним.Якщо виїмки по краю листа неглибокі, лист називається цілісним.Цілісне листя розрізняють за характером виїмок і виступів між ними. Так, якщо виїмки гострі, а виступи заокруглені, лист городчастий(як у шавлії, будри та ін); якщо виїмки клиноподібні, а виступи гострі, трикутні, лист зубчастий(у бука, ліщини та ін.); якщо виступи косокутні та гострі, виходить пильчастийлист (біля груші).
За формою верхівки платівки листя бувають: тупі, гострі, загостреніі гострокінцеві.
За формою основи платівки розрізняють листя клиноподібні, серцеподібні, списоподібні, стрілоподібні та ін.
Крім перерахованих категорій цілісного листа, розрізняють ще лопатеві, роздільніі розсічене листя.Лопатевим називається лист, у якого вирізи по краях пластинки доходять до однієї чверті її ширини (у дуба), а при більшому поглибленні, якщо вирізи досягають більше чверті ширини пластинки, лист називають роздільним(У маку). Лопаті роздільного листа називаються частками. Розсіченимназивають лист, у якого вирізи по краях платівки доходять майже до середньої жилки, утворюючи сегменти платівки.
Роздільні та розсічені листя можуть бути пальчастіі перисті, двічі пальчастіі двічі пір'ястіі т.д.
Що стосується складнихлистя, то серед них розрізняють трійчастосло - ні, пал'чатоскладніі перистоскладні.Якщо складний лист складається з трьох листочків, його називають трійчастоскладним або трійчастим (уконюшини, люцерни, сої, пуерарії та ін.). Якщо черешки листочків прикріплюються до головного черешка як би в одній точці, а самі листочки розходяться радіально, лист називають пал'чатосложнъм(У люпину, коноплі, кінського каштана). Якщо на головному черешку бічні листочки розташовані з обох боків по довжині черешка, лист називають перистоскладні.Якщо такий лист закінчується нагорі непарним одиночним листочком, виходить непарноперистийлист (у еспарцету, білої акації, горобини та ін.). Якщо замість верхнього одиночного листочка розвивається вусик (у вікі), то лист відноситься до непарноперистим.Якщо кінцевого листочка чи вусика немає, лист називають парноперистим(У арахісу, ріжкового дерева). Іноді у непарноперистого листа всі бічні листочки редукуються і залишається тільки кінцевий, силь - непарний листочок, що розвивається, так що лист здається простим, а не перистим (у апельсина).
Аркуш - надзвичайно важливий орган рослини. Аркуш - частина втечі. Основними функціями його є фотосинтез та транспірація. Лист характеризується високою морфологічною пластичністю, різноманітністю форм та великими пристосувальними можливостями. Основа листа може розширюватися у вигляді косих листоподібних утворень - прилистків з кожного боку листка. У деяких випадках вони настільки великі, що відіграють певну роль у фотосинтезі. Прилистки бувають вільними або прирослими до черешка, вони можуть зміщуватися на внутрішній бік листа і тоді їх називають пазушними. Підстави листя можуть бути перетворені у піхву, що оточує стебло і перешкоджають його згинання.
Зовнішня будова листа
Листові платівки різняться за розмірами: від кількох міліметрів до 10-15 метрів і навіть 20 (біля пальм). Тривалість життя листя вбирається у кількох місяців, в деяких — від 1,5 до 15 років. Розмір та форма листя є спадковими ознаками.
Частини листа
Лист - бічний вегетативний орган, що росте від стебла, що має двосторонню симетрію та зону зростання на підставі. Лист зазвичай складається з листової пластинки, черешка (винятком є сидяче листя); для ряду сімейств характерні прилистки. Листя буває просте, що має одну листову пластинку, і складне - з декількома листовими пластинками (листочками).
Листова платівка- Розширена, зазвичай плоска частина листа, що виконує функції фотосинтезу, газообміну, транспірації і в деяких видів - вегетативного розмноження.
Основа листа (листова подушка)- Частина листа, що з'єднує його зі стеблом. Тут знаходиться освітня тканина, що дає ріст листовій платівці та черешку.
Прилистки- парні листоподібні утворення на основі листа. Вони можуть опадати при розгортанні листа або зберігатися. Захищають пазушні бічні бруньки та освітню вставну тканину листа.
Черешок- звужена частина листа, що з'єднує своєю основою листову пластинку зі стеблом. Він виконує найважливіші функції: орієнтує лист по відношенню до світла, є місцем розташування вставної освітньої тканини, рахунок якої росте лист. Крім цього, він має механічне значення для ослаблення ударів по листовій пластинці від дощу, граду, вітру та ін.
Просте та складне листя
Лист може мати одну (простий), кілька або множину листових пластинок. Якщо останні забезпечені зчленуваннями, такий лист називається складним. Завдяки зчленування на загальному черешку листка листочки складного листя опадають поодинці. Однак у деяких рослин складне листя може опадати і цілком.
За формою цілісні листки, розрізняють як лопатеві, роздільні та розсічені.
Лопатевимназиваю лист, у якого вирізи по краях пластинки доходять до однієї чверті його ширини, а при більшому заглибленні, якщо вирізи досягають більше чверті ширини пластинки, лист називається роздільним. Лопаті роздільного листа називають частками.
Розсіченимназивають лист, у якого вирізи по краях пластинки сягають майже середньої жилки, утворюючи сегменти пластинки. Роздільні та розсічені листя можуть бути пальчасті та перисті, двічі пальчасті та двічі перисті і т.д. відповідно до цього розрізняють пальчасто-роздільний лист, перисторозсічений лист; непарно-перисторозсічений листок у картоплі. Він складається з кінцевої частки, кількох пар бічних часточок, між якими розташовуються ще менші часточки.
Якщо платівка подовжена, а частки чи сегменти її трикутні, лист називають стругоподібним(кульбаба); якщо бічні частки нерівновеликі, до основи зменшуються, а кінцева частка велика і округла, виходить ліроподібний лист (редька).
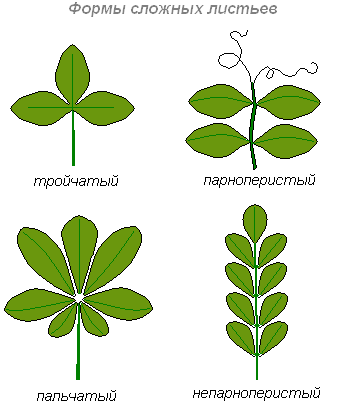
Що стосується складного листя, то серед них розрізняють трійчастоскладне, пальчастоскладне і перистоскладне листя. Якщо складний лист складається з трьох листочків, він називається трійчастоскладним, або трійчастим (клен). Якщо черешки листочків прикріплюються до головного черешка як би в одній точці, а листочки розходяться радіально, лист називається пальчатосложным (люпин). Якщо на головному черешку бічні листочки розташовані по обидва боки по довжині черешка, лист називається перистоскладним.
Якщо такий лист закінчується нагорі непарним одиночним листочком, виходить непарноперистий лист. Якщо ж кінцевого немає, лист називається парноперистим.
Якщо кожен листочок перистосложного листа, своєю чергою, є складним, виходить двічі перистосложный лист.
Форми цілісних листових платівок
Складним листом називають такий, на черешку якого є кілька листових пластин. Вони кріпляться до головного черешка своїми власними черешками, нерідко самостійно, поодинці, опадають, і називаються листочками.
Форми листових пластинок різних рослин відрізняються за контуром, ступенем розчленованості, формою основи і верхівки. Обриси можуть бути овальними, круглими, еліптичними, трикутними та іншими. Листова платівка буває подовженою. Вільний кінець її може бути гострим, тупим, загостреним, гострим. Основа її звужена і відтягнута до стебла, може бути округлим, серцеподібним.

Прикріплення листя до стебла
Листя прикріплюється до втечі довгими, короткими черешками або буває сидячим.
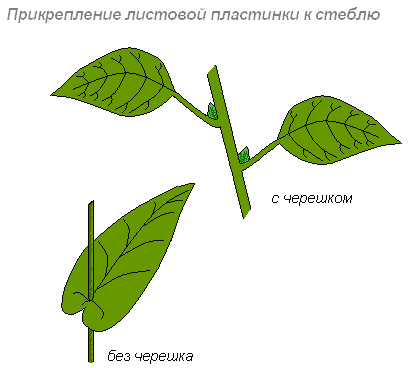
У деяких рослин основа сидячого листа на великому протязі зростається з втечею (нижче лист) або втеча пронизує листову пластинку наскрізь (пронизаний лист).

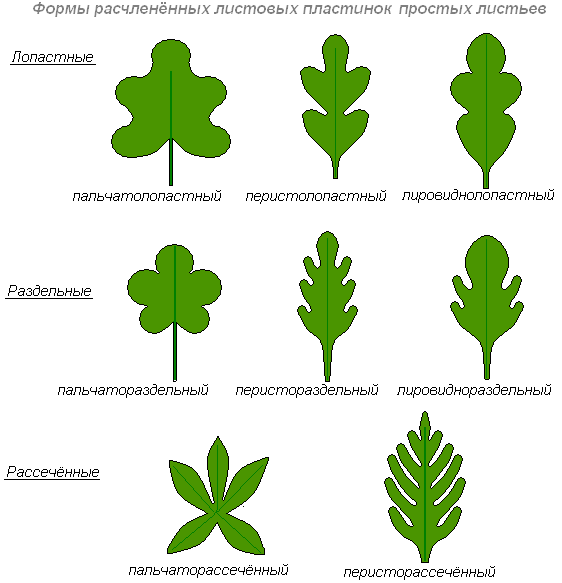
Форма краю листової платівки
Листові пластинки розрізняють за ступенем розсіченості: неглибокі надрізи - зубчасті або пальчасті краї листа, глибокі вирізи - лопатеві, роздільні та розсічені краї.
Якщо краї листової пластинки не мають жодних виїмок, лист називається цілокраїм. Якщо виїмки по краю листа неглибокі, лист називається цілісним.
Лопатевийлист - лист, пластинка якого розчленована на лопаті до 1/3 ширини піваркуша.
Роздільнийлист — лист із пластинкою, розчленованою до ½ ширину піваркуша.
Розсіченийлист - лист, пластинка якого розчленована до головної жилки або до основи листа.
Край листової пластинки - пильчастий (гострі кути).
Край листової платівки - городчастий (округлі виступи).
Край листової пластинки - виїмчастий (округлі виїмки).
Жилкування
На кожному аркуші легко помітити численні жилки, особливо виразні та рельєфні на нижній стороні аркуша.
Жилки- Це провідні пучки, що з'єднують лист зі стеблом. Функції їх — провідна (постачання листя водою та мінеральними солями та виведення з них продуктів асиміляції) та механічна (жилки є опорою для листової паренхіми та захищають листя від розривів). Серед різноманітності жилкування розрізняють листову пластинку з однією головною жилкою, від якої розходяться бічні відгалуження за перистим або пальчатоперистим типом; з декількома головними жилками, що розрізняються товщиною та напрямом розподілу по платівці (дугонервний, паралельний типи). Між описаними типами жилкування існує багато проміжних чи інших форм.
Вихідна частина всіх жилок листової пластинки знаходиться в черешку листа, звідки виходить у багатьох рослин основна, головна жилка, розгалужуючись потім у товщі пластинки. У міру віддалення від головної бічні жилки все витончуються. Найтонші переважно знаходяться на периферії, а також далеко від периферії — посередині ділянок, оточених дрібними жилками.
Існує кілька типів жилкування. У однодольних рослин жилкування буває дугонервним, у якому від стебла чи піхви входить у платівку ряд жилок, дугоподібно спрямованих до вершини пластинки. У більшості злаків має місце паралельно-нервове жилкування. Дугонервне жилкування існує також у деяких дводольних рослин, наприклад подорожника. Однак і вони мають зв'язок між жилками.
У дводольних рослин жилки утворюють сильно розгалужену мережу і відповідно до цього розрізняють жилкування сітчастонервоне, що говорить про краще забезпечення провідними пучками.
Форма основи, верхівки, черешка листа
За формою верхівки пластинки листя бувають тупі, гострі, загострені та гострі.
За формою основи платівки розрізняють листя клиноподібні, серцеподібні, списоподібні, стрілоподібні та ін.

Внутрішня будова листа
Будова шкірки листа
Верхня шкірка (епідерма) - покривна тканина на зверненому боці листа, часто вкрита волосками, кутикулою, воском. Зовні листок має шкірку (покривну тканину), яка захищає його від несприятливих впливів зовнішнього середовища: від висихання, від механічних пошкоджень, від проникнення до внутрішніх тканин хвороботворних мікроорганізмів. Клітини шкірки живі, за розмірами та формою вони різні. Одні з них більші, безбарвні, прозорі та щільно прилягають один до одного, що підвищує захисні якості покривної тканини. Прозорість клітин дозволяє проникати сонячному світлу всередину листа.
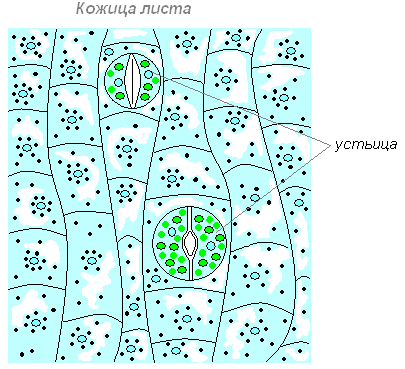
Інші клітини дрібніші, у яких є хлоропласти, які надають їм зелений колір. Ці клітини розташовуються парами і мають здатність змінювати свою форму. При цьому клітини або віддаляються одна від одної, і між ними з'являється щілина або наближаються одна до одної і щілина зникає. Ці клітини назвали замикаючими, а щілину, що виникає між ними, — устьичною. Продихання відкривається, коли замикаючі клітини насичені водою. При відпливі води з замикаючих клітин продихом закривається.
Будова продихи

Через устьичні щілини повітря надходить до внутрішніх клітин листа; через них газоподібні речовини, у тому числі і пари води, виходять з листа назовні. При недостатньому забезпеченні рослини водою (що може статися в суху та спекотну погоду), продихи закриваються. Цим рослини захищають себе від висушення, тому що водяні пари при закритих устьичних щілинах не виходять назовні і зберігаються в міжклітинниках листа. Таким чином, рослини зберігають воду у посушливий період.
Основна тканина листа
Стовпчаста тканина- Основна тканина, клітини якої мають циліндричну форму, щільно прилягають одна до одної і розташовані з верхньої сторони листа (навернутої до світла). Служить для фотосинтезу. Кожна клітина цієї тканини має тонку оболонку, цитоплазму, ядро, хлоропласти, вакуоль. Наявність хлоропластів надає зеленого кольору тканини і всього листа. Клітини, які прилягають до верхньої шкірки листа, витягнуті і розташовані вертикально, називають стовпчастою тканиною.

Губчаста тканина- Основна тканина, клітини якої мають округлу форму, розташовані пухко і між ними утворюються великі міжклітини, також заповнені повітрям. У міжклітинниках основної тканини накопичуються пари води, що надходять сюди із клітин. Служить для фотосинтезу, газообміну та транспірації (випаровування).
Кількість шарів клітин стовпчастої та губчастої тканин залежить від освітлення. У листі, що виросло на світлі, стовпчаста тканина розвинена сильніше, ніж у листя, що виросло в умовах затемнення.

Провідна тканина- Основна тканина листа, пронизана жилками. Жилки - це провідні пучки, так як вони утворені провідними тканинами - лубом та деревиною. По лубу здійснюється передача розчинів цукру з листя всім органам рослини. Рух цукру йде ситоподібними трубками лубу, які утворені живими клітинами. Ці клітини витягнуті в довжину, і в тому місці, де вони стикаються одна з одною короткими сторонами в оболонках, є невеликі отвори. Через отвори в оболонках розчин цукру переходить із однієї клітини до іншої. Ситоподібні трубки пристосовані до передачі органічної речовини на велику відстань. Щільно по всій довжині до бічної стінки ситоподібної трубки прилягають живі клітини менших розмірів. Вони супроводжують клітини трубки, і їх називають клітинами супутницями.
Будова жилок листа

Крім лубу до складу провідного пучка входить і деревина. По судинах листка, як і і докорінно, рухається вода з розчиненими у ній мінеральними речовинами. Воду та мінеральні речовини рослина поглинає з ґрунту корінням. Потім з коріння по судинах деревини ці речовини надходять у надземні органи, у тому числі і до клітин листка.
До складу численних жилок входять волокна. Це довгі клітини з загостреними кінцями і потовщеними здерев'янілими оболонками. Великі жилки листа нерідко оточені механічною тканиною, яка повністю складається з товстостінних клітин - волокон.
Таким чином, по жилках йде передача розчину цукру (органічної речовини) з листа до інших органів рослин, а від кореня – води та мінеральних речовин до листя. З листа розчини рухаються ситоподібними трубками, а до листа — судинами деревини.
Нижня шкірка покривна тканина з нижнього боку листа, зазвичай несе продихання.
Життєдіяльність листа
Зелене листя - органи повітряного харчування. Зелений лист виконує важливу функцію життя рослин — тут утворюються органічні речовини. Будова листа добре відповідає цій функції: він має плоску листову пластинку, а м'якоті листа міститься величезна кількість хлоропластів із зеленим хлорофілом.
Речовини, необхідні для утворення крохмалю в хлоропластах.
Ціль:з'ясуємо, які речовини необхідні освіти крохмалю?
Що робимо:помістимо дві невеликі кімнатні рослини в темне місце. Через два-три дні першу рослину поставимо на шматок скла, а поруч помістимо склянку з розчином їдкого лугу (вона поглине з повітря весь вуглекислий газ), і все це накриємо скляним ковпаком. Для того щоб повітря не надходило до рослини з довкілля, змастимо краю ковпака вазеліном.
Другу рослину також поставимо під ковпак, але тільки поруч із рослиною помістимо склянку з содою (або шматочком мармуру), змоченими розчином соляної кислоти. Внаслідок взаємодії соди (або мармуру) з кислотою виділяється вуглекислий газ. У повітрі під ковпаком другої рослини утворюється багато вуглекислого газу.
Обидві рослини помістимо в однакові умови (світло).
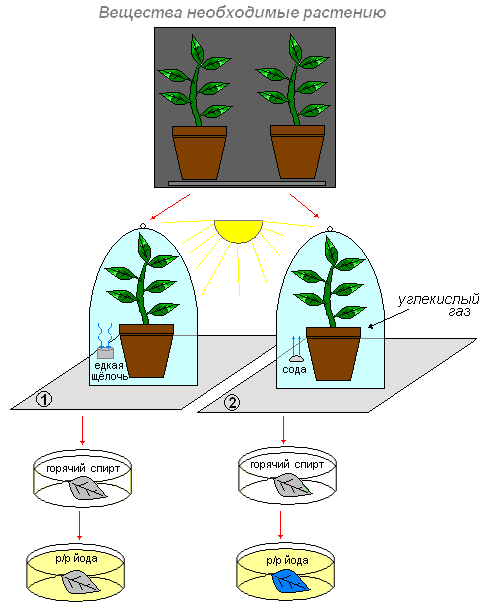
На наступний день візьмемо по листку з кожної рослини та обробимо спочатку гарячим спиртом, промиваємо та діємо розчином йоду.
Що спостерігаємо:у першому випадку фарбування листа не змінилося. Темно-синім став листок тієї рослини, яка була під ковпаком, де був вуглекислий газ.
Висновок:це доводить, що вуглекислий газ необхідний рослині для утворення органічної речовини (крохмаль). Цей газ належить до складу атмосферного повітря. Повітря надходить у листок через устьичні щілини і заповнює простору між клітинами. З міжклітинників вуглекислий газ проникає у всі клітини.
Освіта у листі органічних речовин
Ціль:з'ясувати, у яких клітинах зеленого листа утворюються органічні речовини (крохмаль, цукор).
Що робимо: кімнатна рослинагерань облямована помістимо на три доби у темну шафу (щоб стався відтік поживних речовин з листя). Через три доби виймемо рослину з шафи. Прикріпимо на один із листя конверт із чорного паперу з вирізаним словом «світло» і поставимо рослину на світло або під електричну лампочку. Через 8-10 годин зріжемо лист. Знімемо папір. Опустимо лист у киплячу воду, а потім на кілька хвилин у гарячий спирт (у ньому хлорофіл добре розчиняється). Коли спирт забарвиться у зелений колір, а лист знебарвиться, промиємо його водою та помістимо у слабкий розчин йоду.

Що спостерігаємо:на знебарвленому листі з'являться сині букви (крохмаль синіє від йоду). Літери з'являються на тій частині аркуша, на яку падало світло. Значить, у освітленій частині листа утворився крохмаль. Потрібно звернути увагу на те, що біла смужка по краю листа не забарвилася. Це пояснює те, що в пластидах клітин білої смужки листа герані облямованої немає хлорофілу. Тому крохмаль не виявляється.
Висновок:таким чином, органічні речовини (крохмаль, цукор) утворюються тільки в клітинах з хлоропластами, і для їх утворення необхідне світло.
Спеціальні дослідження вчених показали, що на світлі у хлоропластах утворюється цукор. Потім у результаті перетворень із цукру на хлоропластах утворюється крохмаль. Крохмаль - це органічна речовина, яка у воді не розчиняється.
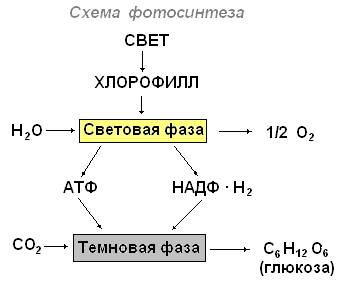
Виділяють світлову та темнову фази фотосинтезу.
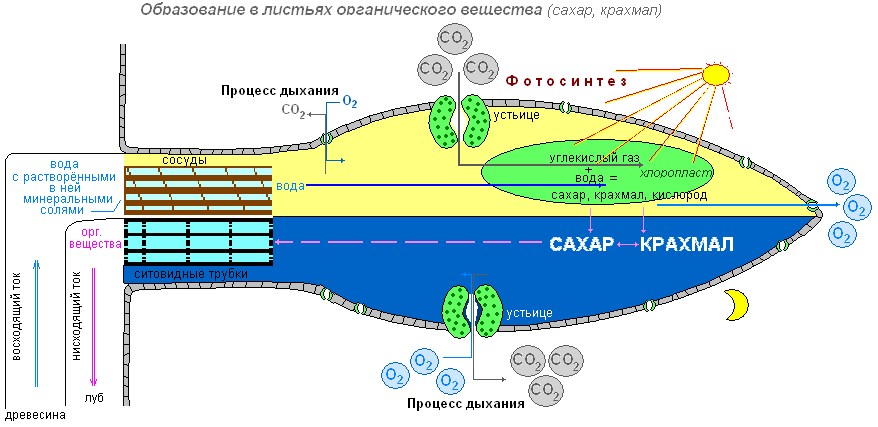
Під час світлової фази фотосинтезу відбувається поглинання світла пігментами, утворення збуджених (активних) молекул, що мають надлишок енергії, йдуть фотохімічні реакції, в яких беруть участь збуджені молекули пігментів. Світлові реакції протікають на мембранах хлоропласту, де знаходиться хлорофіл. Хлорофіл є високоактивною речовиною, що здійснює поглинання світла, первинне запасання енергії та подальше перетворення її в хімічну енергію. У фотосинтезі беруть участь і жовті пігменти каротиноїдів.
Процес фотосинтезу можна у вигляді сумарного рівняння:
6СО 2 + 6Н 2 О = С 6 Н 12 О 6 + 6О 2
Таким чином, суть світлових реакцій у тому, що світлова енергія перетворюється на хімічну.
Темнові реакції фотосинтезу йдуть у матриксі (стромі) хлоропласту за участю ферментів та продуктів світлових реакцій та призводять до синтезу органічних речовин з вуглекислоти та води. Для темнових реакцій не потрібна безпосередня участь світла.
Підсумком темнових реакцій є утворення органічних сполук.
Процес фотосинтезу здійснюється у хлоропластах, у два етапи. У гранах (тілакоїдах) протікають реакції, що викликаються світлом, - світлові, а в стромі - реакції, не пов'язані зі світлом, - темнові, або реакції фіксації вуглецю.
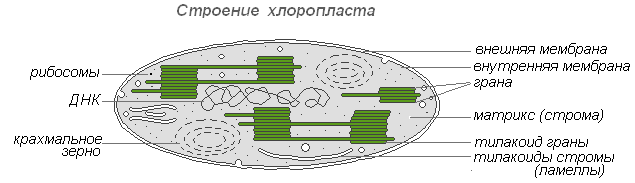
Світлові реакції
1. Світло, потрапляючи на молекули хлорофілу, що знаходяться в мембранах тилакоїдів гран, наводить їх у збуджений стан. В результаті цього електрони сходять зі своїх орбіт і переносяться за допомогою переносників за межі мембрани тилакоїда, де і накопичуються, створюючи негативно заряджене електричне поле.
2. Місце електронів, що вийшли в молекулах хлорофілу займають електрони води ē, так як вода під дією світла піддається фоторозкладу (фотолізу):
Н 2 О↔ОН+Н + ; ВІН‾−ē→ВІН.
Гідроксили ВІН‾, ставши радикалами ВІН, об'єднуються: 4ОН→2Н 2 О+О 2 , утворюючи воду і вільний кисеньщо виділяється в атмосферу.
3. Протони Н+ не проникають через мембрану тилакоїда і накопичуються всередині, використовуючи позитивно заряджене електричне поле, що призводить до збільшення різниці потенціалів з обох боків мембрани.
4. При досягненні критичної різниці потенціалів (200 мВ) протони Н + спрямовуються протонним каналом в ферменті АТФ-синтетазу, вбудованому в мембрану тилакоїда, назовні. На виході з протонного каналу створюється високий рівень енергії, що йде синтез АТФ (АДФ+Ф→АТФ) . Молекули АТФ, що утворилися, переходять у строму, де беруть участь у реакціях фіксації вуглецю.
5. Протони Н + , що вийшли на поверхню мембрани тилакоїда, з'єднуються з електронами ē, утворюючи атомарний водень Н, який йде на відновлення переносників НАДФ + : 2ē+2Н + = НАДФ + → НАДФ∙Н 2 (переносник з приєднаним воднем; ).
Таким чином, активований світловий енергією електрон хлорофілу використовується для приєднання водню до переносника. НАДФ∙Н2 перетворюється на строму хлоропласту, де бере участь у реакціях фіксації вуглецю.
Реакції фіксації вуглецю (темнові реакції)
Здійснюється у стромі хлоропласту, куди надходять АТФ, НАДФ∙Н 2 від тилакоїдів гран та СО 2 з повітря. Крім того, там постійно перебувають п'ятивуглецеві сполуки — пентози С 5 , які утворюються в циклі Кальвіна (цикл фіксації СО 2). Спрощено цей цикл можна так:
1. До пентози 5 приєднується 2 , в результаті чого з'являється нестійке шестикутне з'єднання 6 , яке розщеплюється на дві тривуглецеві групи 2С 3 - тріози.
2. Кожна із тріоз 2С 3 приймає по одній фосфатній групі від двох АТФ, що збагачує молекули енергією.
3. Кожна з тріоз 2С 3 приєднує по одному атому водню від двох НАДФ Н2.
4. Після чого одні тріози об'єднуються, утворюючи вуглеводи 2С 3 → 6 → 6 Н 12 О 6 (глюкоза).
5. Інші тріози об'єднуються, утворюючи пентози 5С 3 →3С 5 і знову включаються в цикл фіксації СО 2 .
Сумарна реакція фотосинтезу:
6СО 2 +6Н 2 Про хлорофіл енергія світла →С 6 Н 12 Про 6 +6О 2
Окрім вуглекислого газу в освіті крохмалю бере участь вода. Її рослина отримує із ґрунту. Коріння поглинає воду, яка по судинах пучків, що проводять, піднімається в стебло і далі в листя. А вже у клітках зеленого листа, у хлоропластах, з вуглекислого газу та води за наявності світла утворюється органічна речовина.
Що відбувається з органічними речовинами, утвореними у хлоропластах?
Крохмаль, що утворився в хлоропластах, під впливом особливих речовин перетворюється на розчинний цукор, який надходить до тканин усіх органів рослини. У клітинах деяких тканин цукор може знову перетворитися на крохмаль. Запасний крохмаль накопичується у безбарвних пластидах.
З цукрів, що утворилися при фотосинтезі, а також мінеральних солей, поглинених корінням із ґрунту, рослина створює речовини, які йому необхідні: білки, жири та багато інших білків, жирів та багато інших.
Частина органічних речовин, синтезованих у листі, витрачається на зростання та харчування рослини. Інша частина відкладається у запас. У однорічних рослин запасні речовини відкладаються у насінні, плодах. У дворічних на першому році життя вони накопичуються в вегетативні органи. У багаторічних травречовини запасаються в підземних органах, а в дерев і чагарників - в серцевині, основної тканини кори та деревини. Крім того, у них на певному році життя органічні речовини починають запасатися також у плодах та насінні.
Типи живлення рослини (мінеральне, повітряне)
У живих клітинах рослини постійно відбувається обмін речовин та енергії. Одні речовини поглинаються та використовуються рослиною, інші виділяються у навколишнє середовище. З найпростіших речовин утворюються складні. Складні органічні речовини розщеплюються прості. Рослини накопичує енергію, а процесі фотосинтезу і звільняє її при диханні, використовуючи цю енергію реалізації різних процесів життєдіяльності.
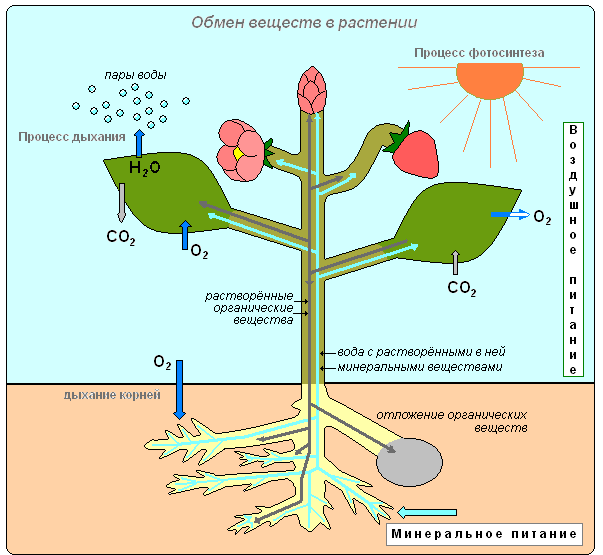
Газообмін
Листя завдяки роботі продихів здійснюють і таку важливу функцію, як газообмін між рослиною та атмосферою. Через продихи лист з атмосферним повітрям надходять вуглекислий газ і кисень. Кисень використовується при диханні, вуглекислий газ необхідний рослині для утворення органічних речовин. Через продихи в повітря виділяється кисень, який утворився в процесі фотосинтезу. Видаляється і вуглекислий газ, що у рослини у процесі дихання. Фотосинтез здійснюється лише світла, а дихання світла й у темряві, тобто. постійно. Дихання у всіх живих клітинах органів рослини відбувається безперервно. Як і тварини, рослини гинуть із припиненням дихання.
У природі відбувається обмін речовин між живим організмом та навколишнім середовищем. Поглинання рослиною одних речовин із зовнішнього середовища супроводжується виділенням інших. Елодея, будучи водяною рослиною, використовує для харчування вуглекислий газ, розчинений у воді.
Ціль:з'ясуємо, яку ж речовину виділяє елодея у довкілля при фотосинтезі?
Що робимо:стебла гілочок підріжемо під водою (вода кип'ячена) біля основи і прикриємо скляною лійкою. Пробірку до країв заповнену водою поміщаємо на трубку воронки. Це зробити у двох варіантах. Одну ємність поставити в темне місце, а іншу - виставити на яскраве сонячне або штучне світло.
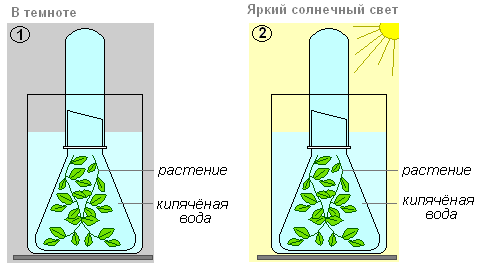
У третю і четверту ємності додати вуглекислий газ (додати невелику кількість питної соди або можна подихати в трубочку) і один поставити в темряву інший на сонячне світло.

Що спостерігаємо:через деякий час у четвертому варіанті (посудина, що стоїть на яскравому сонячному світлі) починають виділятися бульбашки. Цей газ витісняє воду з пробірки, її рівень у пробірці витісняється.
Що робимо:коли вода буде витіснена газом повністю, необхідно обережно зняти пробірку з лійки. Щільно закрити отвір великим пальцем лівої руки, а правою швидко внести в пробірку лучинку, що тліє.
Що спостерігаємо:лучинка загоряється яскравим полум'ям. Подивившись на рослини, які помістили у темряву, побачимо, що бульбашки газу з елодеї не виділяються і пробірка залишилася заповнена водою. Те саме з пробірками в першому і другому варіанті.
Висновок:звідси випливає, що газ, який виділила елодея, — кисень. Таким чином, рослина виділяє кисень лише тоді, коли є всі умови для фотосинтезу – вода, вуглекислий газ, світло.
Випаровування води листям (транспірація)
Процес випаровування води листям у рослин регулюється відкриванням та закриванням продихів. Закриваючи продихи, рослина захищає себе від втрати води. Відкриття та закривання продихів знаходиться під впливом факторів зовнішнього та внутрішнього середовища, в першу чергу температури та інтенсивності сонячного світла.
Листя рослин містить багато води. Вона надходить за провідною системою від коріння. Усередині листа вода просувається по стінках клітин і міжклітинникам до продихів, через які йде у вигляді пари (випаровується). Цей процес легко перевірити, якщо виконати нескладний пристрій, як показано на малюнку.

Випаровування води рослиною називається транспірацією. Воду випаровує поверхню листка рослини, особливо інтенсивно - поверхню листка. Розрізняють кутикулярну транспірацію (випаровування всією поверхнею рослини) і устьичну (випаровування через устячка). Біологічне значення транспірації полягає в тому, що вона є засобом пересування води та різних речовин по рослині (присмоктує дію), сприяє надходженню вуглекислого газу всередину листа, вуглецевому живленню рослин, захищає листя від перегріву.
Інтенсивність випаровування води листям залежить від:
- біологічних особливостей рослин;
- умов зростання (рослини посушливих місцевостей випаровують мало води, вологих - значно більше; тіньові рослини випаровують води менше, ніж світлові; багато води рослини випаровують у спеку, значно менше - у хмарну погоду);
- освітлення (розсіяне світло зменшує транспірацію на 30-40%);
- вміст води в клітинах листа;
- осмотичного тиску клітинного соку;
- температури ґрунту, повітря та тіла рослини;
- вологості повітря та швидкості вітру.
Найбільша кількість води випаровується у деяких видів деревних порід через листові рубці (рубець, що залишається опалим листям на стеблі), які виявляються найбільш вразливими місцями на дереві.
Взаємозв'язок процесів дихання та фотосинтезу
Весь процес дихання протікає у клітинах рослинного організму. Він складається з двох етапів, у ході яких органічні речовини розщеплюються на вуглекислий газ та воду. На першому етапі за участю спеціальних білків (ферментів) відбувається розпад молекул глюкози на простіші органічні сполуки і виділяється трохи енергії. Цей етап дихального процесу відбувається у цитоплазмі клітин.
На другому етапі прості органічні речовини, що утворилися на першому етапі, під дією кисню розпадаються на вуглекислий газ та воду. У цьому вивільняється багато енергії. Другий етап дихального процесу протікає лише за участю кисню та у спеціальних тільцях клітини.
Поглинені речовини у процесі перетворень у клітинах і тканинах стають речовинами, у тому числі рослина будує своє тіло. Усі перетворення речовин, що відбувається в організмі, завжди супроводжуються споживанням енергії. Зелена рослина як автотрофний організм, поглинаючи світлову енергію Сонця, накопичує її в органічних сполуках. У процесі дихання при розщепленні органічних речовин ця енергія вивільняється та використовується рослиною для процесів життєдіяльності, що відбуваються у клітинах.
Обидва процеси - фотосинтез і дихання - йдуть шляхом послідовних численних хімічних реакцій, у яких одні речовини перетворюються на інші.
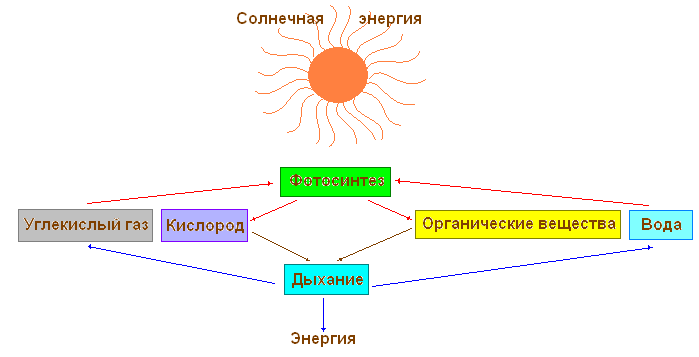
Так, у процесі фотосинтезу з вуглекислого газу та води, отриманих рослиною з навколишнього середовища, утворюються цукру, які потім перетворюються на крохмаль, клітковину або білки, жири та вітаміни – речовини, необхідні рослині для харчування та запасання енергії. У процесі дихання, навпаки, відбувається розщеплення створених у процесі фотосинтезу органічних речовин на неорганічні сполуки – вуглекислий газ та воду. При цьому рослина отримує енергію, що вивільняється. Ці перетворення речовин в організмі називають обміном речовин. Обмін речовин - одна з найважливіших ознак життя: із припиненням обміну речовин припиняється життя рослини.
Вплив факторів середовища на будову аркуша
Листя рослин вологих місць, як правило, велике з великою кількістю продихів. З поверхні цього листя випаровується багато вологи.
Листя рослин посушливих місць невеликі за розміром і мають пристрої, що зменшують випаровування. Це густе опушення, восковий наліт, відносно невелика кількість продихів та ін. У деяких рослин листя м'яке і соковите. Вони запасається вода.

Листя тіньовитривалих рослин має всього два-три шари округлих, нещільно прилеглих один до одного клітин. Великі хлоропласти розташовані так, що не затіняють один одного. Тіньове листя, як правило, більш тонке і має більш темне зелене забарвлення, оскільки містить більше хлорофілу.
У рослин відкритих місць м'якуш листа налічує кілька шарів, щільно прилеглих один до одного стовпчастих клітин. У них міститься менше хлорофілу, тому світлове листя має більш світле забарвлення. Те й інше листя іноді можна зустріти і в кроні одного і того ж дерева.
Захист від зневоднення
Зовнішня стінка кожної клітини шкірки листа не тільки потовщена, а й захищена кутикулою, яка погано пропускає воду. Захисні властивості шкірки значно підвищуються при утворенні волосків, що відбивають сонячні промені. Завдяки цьому нагрівання листа знижується. Все це обмежує можливість випаровування води із поверхні листа. При нестачі води закривається устьична щілина і пара не виходить назовні, накопичуючись у міжклітинниках, що призводить до припинення випаровування з поверхні листа. Рослини жарких і сухих місць проживання мають невелику платівку. Чим менша поверхня листа, тим менша небезпека зайвої втрати води.
Видозміни листя
У процесі пристосування до умов навколишнього середовища листя у деяких рослин видозмінилися тому, що стали відігравати роль не властиву типовим листям. У барбариса частина листя видозмінилася на колючки.

Старіння листя та листопад
Листопад передує старіння листя. Це означає, що у всіх клітинах знижується інтенсивність життєвих процесів – фотосинтезу, дихання. Зменшується вміст вже наявних у клітинах важливих для рослини речовин і скорочується надходження нових, у тому числі води. Розпад речовин переважає їх утворення. У клітинах накопичуються непотрібні, і навіть шкідливі продукти, їх називають кінцевими продуктами обміну речовин Ці речовини видаляються з рослини під час скидання листя. Найбільш цінні сполуки по провідних тканин відтікають з листя в інші органи рослини, де відкладаються в клітинах запасних тканин або відразу використовується організмом для харчування.
У більшості дерев і чагарників у період старіння листя змінює забарвлення і стає жовтим або багряним. Це тому, що хлорофіл руйнується. Але крім нього в пластидах (хлоропластах) є речовини жовтого та оранжевого кольору. Влітку вони були ніби замасковані хлорофілом і пластиди мали зелений колір. Крім того, у вакуолях накопичуються інші барвники жовтого або червоно-малинового кольору. Разом із пігментами пластид вони визначають забарвлення осіннього листя. У деяких рослин листя зберігає зелений колір до відмирання.
Ще до того, як з втечі впаде лист, у його основі на кордоні зі стеблом формується шар пробки. Зовні від нього утворюється шар. Згодом клітини цього шару одягаються одна від одної, оскільки ослизняется і руйнується міжклітинна речовина, яка їх з'єднувала, а іноді й оболонки клітин. Лист відокремлюється від стебла. Однак деякий час він ще зберігається на втечі завдяки пучкам, що проводять, між листом і стеблом. Але настає момент порушення цього зв'язку. Рубець на місці листа, що відокремився, покритий захисною тканиною, пробкою.

Як тільки листя досягає граничних розмірів, починаються процеси старіння, що ведуть зрештою до відмирання листка — його пожовтіння або почервоніння, пов'язане з руйнуванням хлорофілу, накопиченням каротиноїдів і антоціанів. У міру старіння листка знижується також інтенсивність фотосинтезу та дихання, деградують хлоропласти, накопичуються деякі солі (кристали оксалати кальцію), з листка відтікають пластичні речовини (вуглеводи, амінокислоти).
У процесі старіння листа біля його основи у дводольних деревних рослинформується так званий відокремлювальний шар, який складається з паренхіми, що легко розшаровується. За цим шаром листок і відокремлюється від стебла, причому на поверхні майбутнього листового рубцязаздалегідь утворюється захисний шар коркової тканини.

На листовому рубці помітні у вигляді точок поперечні перерізи листового сліду. Скульптура листового рубця різна і є характерною ознакоюдля систематики лепідофіт.
У однодольних і трав'янистих дводольних відокремлювальний шар, як правило, не утворюється, листок відмирає і руйнується поступово, залишаючись на стеблі.
У листопадних рослин опадіння листя на зиму має пристосувальне значення: скидаючи листя, рослини різко зменшують випарову поверхню, захищаються від можливих поломокпід вагою снігу. У вічнозелених рослинмасовий листопад приурочений зазвичай до початку зростання нових пагонів із нирок і тому відбувається не восени, а навесні.
Осінній листопад у лісі має важливе біологічне значення. Опале листя — гарне органічне та мінеральне добриво. Щороку на їх листяних лісах опале листя є матеріалом для мінералізації, що виробляється грунтовими бактеріями і грибами. Крім того, опале листя стратифікує насіння, що опало до листопада, оберігає коріння від вимерзання, перешкоджає розвитку мохового покриву і т.д. деякі види дерев скидають не тільки листя, а й однорічні пагони.
Математична модель кінетики росту рослин
Ковпак Євген Петрович,
доктор фізико-математичних наук,
Стовпова Марія Володимирівна,
аспірант.
Санкт-Петербурзький державний університет.
Mathematical Model of Plant Growth Kinetics
Maria Stolbová
doctoral student, St. Petersburg State University.
Evgenii Kolpak
D.Sc, St. Petersburg State University.
У роботі наводяться результати досліджень із вивчення кінетики росту рослин. На основі експериментальних даних запропоновано математичну модель зміни лінійних розмірів рослин, що є завданням Коші для звичайного диференціального рівняння.
Ключові слова:математичне моделювання, морфогенез, кінетика зростання.
Цей paper describes results of study in the kinetics of plant rowth and offers thematematical model of changes in their dimensions базується на experimental data obtained. Model є Cauchy проблемою для звичайних різних еквівалентів.
Keywords:математичне моделювання, морфогенез, зростання кінетики.
Динаміка зростання рослин вперше, мабуть, описана в роботах Сакса (1832 - 1897) - лінійний розмір рослин у часі в його експериментах змінювався за "логістичної" залежності. На сьогоднішній день численні експериментальні дані, опубліковані в літературних джерелах, з різним ступенем точності узгоджуються з характером змін, як лінійних розмірів, так і сумарної біомаси рослин. Однак, для опису зміни «параметра», що характеризує як зростання окремої рослини, так і накопичення їх загальної біомаси, пропонуються різні апроксимуючі залежності такі, як експоненційна, лінійна, параболічна та інші, що не враховують внутрішні біологічні процеси, що зумовлюють зростання рослин та зовнішні впливи , такі як додаткове харчування, температурні зміни, антропогенний вплив. Діяльність пропонується математична модель зростання окремої рослини, розроблена з урахуванням авторських експериментальних даних.
Аналіз кінетики росту рослин проводився таких рослинах як гречка, просо, момордика, лагенарія, лаванда, чуфа, тюльпан та інших.Дослідження проводилися з 2000 по 2012 рік на навчально-дослідній ділянці Кінгісепської станції юних натуралістів та в теплицях ЗАТ «Райдуга» Кінгісепського району. В експериментах брали участь Столбова М.В., Мерзлякова С.М., Лихачова Н.В.
Всерослини (табл. 1), крім тюльпанів, вирощувалися у літній період у природних умовах з 2000 по 2012 рр. Для тюльпанів проводилася вигонка в зимовий період в умовах, за яких регулювалася температура ґрунту та повітря. На вирощування кожного сорту виділялася площа 10 кв.м. Деякі рослини вимагали передпосівного обробітку насіння, вирощування розсади, підготовки грунту з його дезінфекцією розчином марганцево-кислого калію. На постійне місце висаджувалися (висівалися) тоді, коли минула загроза повернення заморозків. Додаткове харчуваннярослинам давали у вигляді підживлення складним мінеральним добривом. Прополку і полив проводили в міру потреби. У процесі росту рослин проводилися виміри висоти рослин механічним способом протягом усього вегетаційного. Висота рослин вимірювалася за допомогою лінійки приблизно 1 раз на 7-10 днів. Температура вимірювалася щодня.
На рис. 1 наведено експериментальні дані (позначені зірочками) для гречки 1. Аналогічні залежності (узгоджується з даними, опублікованими в) отримані і для інших рослин (табл. 1) за весь період проведення експерименту. Максимальна висота рослин змінювалася від 17 см до 110 см. Час зростання від 80 до 110 днів.
Мал. 1. Залежність "висота рослини - час" для гречки 1.
Усі експериментальні дані з кінетики зростання близькі до логістичної залежності. Тобто, для опису динаміки росту рослин можна використовувати рівняння:

де – час (дні), − поточна висота рослини (см), − теоретична максимальна висота (см), яку може досягти рослина після закінчення зростання, − константа (питома швидкість росту, розмірність – 1/день). Рішенням даного рівняння є функція (− початкова висота рослини):
 .
.
Ця залежність використовувалася для опису отриманих експериментальних даних. До онстанти і підбиралися із застосуванням методу найменших квадратів. Результати обробки експериментів (константа) для деяких рослин наведено у табл. 1. Як випливає з отриманих результатів, константи досліджуваних рослин змінювалися в діапазоні 0.06 – 0.15. Похибка їх визначення за три роки вимірів за всіма культурами становила трохи більше 5 %.
Таблиця 1.
Вирощувані рослини та розрахункові значення питомих швидкостей зростання.
|
Назва рослини |
Питома швидкість зростання () |
Назва рослини |
Питома швидкість зростання () |
|
Гречка 1 |
0.15 |
Просо казанське 176 |
0.07 |
|
Гречка 2 |
0.17 |
Тюльпан Denise |
0.06 |
|
Просо вільне |
0.09 |
Тюльпан Denmark |
0.09 |
|
Просо швидке |
0.08 |
Тюльпан Escape |
0.09 |
Одним із найважливіших факторів, що впливають на зростання рослин, є температура. Як випливає з наших експериментальних даних, зміну температури в часі протягом вегетаційного періоду можна описати функцією
де – мінімальна температура за вегетаційний період, а – максимальна – частота зміни максимальних значень температури.
Рослини, з якими проводився експеримент, розвиваються, якщо температура повітря змінюється в діапазоні від (10°С в експерименті) (30°С в експерименті). Якщо вважати, що швидкість зростання максимальна за температури ![]() тоді питома швидкість росту рослини буде пропорційна функції
тоді питома швидкість росту рослини буде пропорційна функції
 якщо,
якщо,
якщо або ,
де − значення температури у поточний момент часу.
Ця функція температури набуває нульових значень приі і досягає екстремуму рівного 1 при . Аналогічний підхід обліку впливу температури зростання рослин використовувався в .
Рівняння для швидкості росту рослин з урахуванням введеного температурного режиму набуде вигляду:
![]() , якщо ,
, якщо ,
Якщо або .
У цій моделі передбачається, що рослина не гине при «порушенні» температурного режиму, а лише припиняється його зростання. Чисельне рішення диференціальних рівнянь та обробку експериментальних даних зручніше реалізовувати серед програмування математичного пакета Matlab , має набір необхідних вбудованих функцій.
Таким чином, облік температурного режиму може точніше описати експериментальні дані та пояснити відхилення експериментальних даних від логістичної залежності більш «біологічно» обґрунтованої, ніж поліномінальні функції.
Література
1. Баранов В.Д., Устименко Г.В. Світ культурних рослин. М.: Думка, 1994. 232 с.
2. Винокурова Р.І., Сілкіна О.В. Ростові характеристики хвої дерев ялиці сибірської (Abies Sibiricf L.) та ялини звичайної (Picea Abies L.) // Вісник МарДТУ. 2008. № 2. С. 40 - 50.
3. Горбунова Є.А., Ковпак Є.П. Математичні моделі одиночної популяції// Вест. С.-Петерб. ун-ту. Сер. 10: Прикладна математика, інформатика та процеси управління. 2012. Вип. 4. З. 18 – 30.
4. Зайцев Г.М. Математична статистика в експериментальній ботаніці. - М.: Наука, 1984. 424 с.
5. Звягінцев А.Ю. Морське обростання у північно-західній частині Тихого океану. Владивосток: Дальнаука, 2005. 432 с.
6. Злобін Ю.А. Екологія популяції рослин: сучасний стан. Суми: Університетська книга 209. 263 с.
7. Ковпак Є.П. MatLab: методи обчислень/Санкт-Петербурзький держ. ун-т. Санкт-Петербург, 2007. 100 с.
8. Кузнєцов В.І., Козлов Н.І., Хом'яков П.М. Математичне моделювання еволюції лісу з метою управління лісовим господарством. М: Ленад. 2005. 232 с.
9. Медведєв С.С. Фізіологія рослин: Підручник. - Спб.: Вид-во С.-Петерб. ун-ту, 2004. 336 с.
10. Назарова С.А., Генельт-Яновський Є.А., Максимович Н.В. Лінійне зростання Macoma Balthica в осушеній зоні мурманського узбережжя Баренцева моря // Вісник СПбГУ. Сер. 3. 2010. Вип. 4. С. 35 - 43.
11. Разін Г.С., Рогозін М.В. Про хід зростання деревостанів. Догматизм у лісовій таксації // Вісник Пермського університету. Біологія 2009. Вип. 10 (36). с. 9 – 38.
12. Раїлкін А.І. Колонізація твердих тіл бентосними організмами. - СПб.: Вид-во С.-Петерб. ун-ту, 2008. 427 с.
13. Суханова Є.С., Кочкін Д.В., Титова М.В., Носов А.М. Ростові та біосинтетичні характеристики різних штамів культур клітин рослин роду Polyscias // Вісник ПДТУ. 2012. № 2. С. 57 - 66.
14. Уорінг Ф., Філіпс І.Ф. Зростання рослини та диференціювання. М: Світ. 1984. 512 с.
15. Усольцев В.А., Воробейчик О.Л., Бергман Біологічна продуктивність лісів Уралу за умов техногенного забруднення: дослідження системи зв'язків та закономірностей. Єкатеринбург: УГЛТУ. 2012. - 366 с.
16. Hewatt WG. Ecological succession в The Mytilus californianus habitat asObserved in Monterey Bay // Cal. Ecol. 1935. V. 16. P. 244-251.
17. Prisman T.I., Slyusar N.A. Mathematical model of seasonal growth of halophytic plant community with account of environmental factors: International meeting of soil fertility land management and agro climatology. Turkey, 2008. P. 43-51.
18. Urban H.J. Modeling growth of different developmental stages in bivalves // Mar. Ecol. Прог. Ser. 2002. Vol. 238. P. 109-114.
19. Wahl M. Living зарахований: Aufwuchs, fouling, epibiosis // Fouling Organisms в Indian Ocean: Біологія і Control Technology (Nagabhushanam R., Thompson M.F., Eds.). New Delhi: Oxford та IBH Publ. Co, 1997. P. 31-83.
20. Wahl M. Marine epibiosis. I. Fouling and antifouling: деякі основні аспекти // Mar. Ecol. Progr. Ser. 1989. Vol. 58, N 1-2. P. 175-189.
21. Wahl M., Hoppe K. Interactions між substratum rugosity, colonization density and periwinkle grazing efficiency // Mar. Ecol. Прог. Ser. 2002. Vol. 225. P. 239-249.
Аркуш - Це бічна спеціалізована частина втечі.
Основні та додаткові функції листа
Основні: функції фотосинтезу, газообміну та випаровування води (транспірація).
Додаткові: вегетативне розмноження, запасання речовин, захисна (колючки), опорна (усики), поживна (у комахоїдних рослин), видалення деяких продуктів обміну речовин (з опаданням листя). Листя росте переважно до певних розмірів за рахунок крайовий меристеми . Зростання їх обмежене (на відміну від стебла та кореня) лише до певних розмірів. Розміри – різні, від кількох міліметрів до кількох метрів (10 і більше).
Термін життя різний. У однорічних рослин листя відмирає разом з іншими частинами тіла. Багаторічні рослиниможуть замінювати листя поступово, протягом вегетаційного періоду або протягом життя – вічнозелені рослини (лавр благородний, фікус, монстера, брусниця, верес, барвінок, лавровишня, пальма тощо). Опадіння листя у несприятливі пори року отримало назву – листопад . Рослини, у яких спостерігається листопад, називаються листопадними (Яблуня, клен, тополя тощо).

Аркуш складається з листової платівки і черешка . Листова платівка плоска. На листовій пластинці можна виділити основу, кінчик та краї. У нижній частині черешка розташоване потовщене заснування листа. У листовій платівці розгалужуються жилки - судинно-волокнисті пучки. Виділяють центральну та бічні жилки. Черешок обертає платівку для кращого уловлювання променів світла. Аркуш опадає разом із черешком. Листя, що має черешок, називаються черешковими . Черешки бувають короткими або довгими. Листя, що не має черешка, називаються сидячими (Наприклад, у кукурудзи, пшениці, наперстянки). Якщо нижня частина листової пластинки охоплює стебло у вигляді трубки або жолобка, то листове утворюється піхву (У деяких злаків, осок, парасолькових). Воно захищає стебло від пошкоджень. Втеча може пронизувати листову пластинку. пронизаний лист .


Форми черешка
На поперечному зрізі черешки можуть мати форму: циліндричну, ребристу, плоску, крилату, жолобчасту тощо.
Деякі рослини (розоцвіті, бобові тощо), крім платівки та черешка, мають особливі вирости – прилистки . Вони прикривають бічні бруньки та захищають їх від пошкоджень. Прилистки можуть мати вигляд маленьких листків, плівок, колючок, лусочок. У деяких випадках бувають дуже великими та відіграють важливу роль у фотосинтезі. Бувають вільними або прирослими до черешка.

Жилки поєднують листок зі стеблом. Це – судинно-волокнисті пучки. Їхні функції: провідна та механічна (жилки служать опорою, захищають від розриву листя). Розташування, розгалуження жилок листової платівки називається жилкуванням . Розрізняють жилкування з однієї головної жилки, від якої розходяться бічні відгалуження – сітчасте, перисте (черемха тощо), пальчасте (клен татарський тощо), або з кількома головними жилками, які йдуть майже паралельно одна одній – дугове (подорожник, конвалія) та паралельне (пшениця, жито) жилкування. Крім того, є багато перехідних типів жилкування.
Для більшості дводольних характерно перисте, пальчасте, сітчасте жилкування, для однодольних – паралельне та дугове.
Листя з прямими жилками переважно цілокраї.
Різноманітність листя за зовнішньою будовою

По листовій платівці:
Розрізняють листя просте і складне.
Просте листя
Прості листя має одну листову пластинку з черешком, яка може бути цілісною або розчленованою. Просте листя опадає під час листопада повністю. Вони поділяються на листя з цільною та розчленованою листовою платівкою. Листя з цільною листовою пластинкою називаються цільними .
Форми листової пластинки відрізняються загальним контуром, формою верхівки та основи. Контур листової пластинки може бути овальним (акація), серцеподібним (липа), голчастим (хвойні), яйцеподібним (груша), стрілоподібним (стрілолист) тощо.
Кінчик (верхівка) листової пластинки буває гострим, тупим, притупленим, загостреним, виїмчастим, усикоподібним тощо.
Основа листової пластинки може бути округлим, серцеподібним, стрілоподібним, списоподібним, клиноподібним, нерівностороннім і т.п.
Край листової пластинки може бути цілокраї або з виїмками (не досягають ширини пластинки). За формами виїмок по краю листової пластинки розрізняють листя зубчасті (зубці мають рівні сторони – ліщина, бук тощо), пильчасті (одна сторона зубця довша за іншу – груша), борідчасті (виїмки гострі, опуклості тупі – шавлія) та ін.
Складне листя
Складні листя має загальний черешок (рахіс). До нього кріпляться прості листочки. Кожен із листочків може опадати самостійно. Складне листя поділяється на трійчасте, пальчасте і перисте. Складні трійчасті листя (конюшина) мають три листочки, які короткими черешками кріпляться до загального черешка. Пальчасті складні листя подібне до будови попереднім, але кількість листочків більше трьох. Перистоскладні листя складаються з листочків, розташованих по всій довжині рахісу. Бувають парноперистоскладні та непарноперистоскладні. Парноперистоскладні листя (горох посівний) складаються з простих листочків, які попарно розташовані на черешку. Непарноперистоскладні листя (шипшина, горобина) закінчуються одним непарним листочком.
За способом членування
Листя поділяють на:
1) лопатеві якщо членування листової пластинки доходить до 1/3 всієї її поверхні; виступаючі частини називають лопатями ;
2) роздільні якщо членування листової пластинки доходить до 2/3 всієї її поверхні; виступаючі частини називають частками ;
3) розсічені якщо ступінь членування доходить до центральної жилки; виступаючі частини називають сегментами .
Листорозташування
Це розташування певному порядку листя на стеблі. Листорозташування – це спадковий ознака, але під час розвитку рослини при пристосуванні до умов освітлення може змінюватися (наприклад, у нижній частині листорозташування протилежне, у верхній – чергове). Розрізняють три види листорозташування: спіральне, або чергове, супротивне і кільчасте.

Спіральне
Властивості більшості рослин (яблуня, береза, шипшина, пшениця). При цьому від вузла відходить лише один аркуш. Розташоване листя на стеблі по спіралі.
Протилежне
У кожному вузлі два листи сидять один навпроти іншого (бузок, клен, м'ята, шавлія, кропива, калина тощо). У більшості випадків листя двох сусідніх пар відходить у двох взаємно протилежних площинах, не затінюючи один одного.
Кільчасте
Від вузла відходить більше двох листків (елодея, вороне око, олеандр тощо).
Форма, розмір та розташування листя пристосовані до умов освітлення. Взаємне розташування листя нагадує мозаїку, якщо подивитися на рослину зверху у напрямку світла (у граба, в'яза, клена та ін.). Таке розташування називається листової мозаїкою . При цьому листя не затінюють один одного і використовують світло ефективно.

Зовні лист покритий переважно одношаровим, іноді багатошаровим епідермісом (шкіркою). Він складається з живих клітин, більшість з яких позбавлені хлорофілу. Крізь них сонячні промені легко потрапляють до нижчих верств клітин листа. У більшості рослин шкірка виділяє та створює зовні тонку плівку з жироподібних речовин – кутикулу, яка майже не пропускає воду. На поверхні деяких клітин шкіри можуть бути волоски, шипики, які захищають листок від пошкоджень, перегріву, надмірного випаровування води. У рослин, які ростуть на суші, на нижній стороні листка в епідермісі є продихи (у вологих місцях (капуста) - продихи з обох сторін листя; у водяних рослин (водяна лілія), листя яких плаває на поверхні, - на верхній стороні; рослин, які занурені повністю у воду, продихів немає). Функції продихів: регуляція газообміну та транспірації (випаровування води листям). У середньому на 1 квадратний міліметр поверхні припадає 100-300 продихів. Чим вище лист розташований на стеблі, тим більше продихів на одиницю поверхні.
Між верхнім та зовнішнім шарами епідермісу розташовані клітини основної тканини – асиміляційної паренхіми. У більшості видів покритонасінних розрізняють два види клітин цієї тканини: стовпчасту (палісадну) і губчасту (пухку) хлорофілоносні паренхіми. Разом вони становлять мезофіл листа. Під верхньою шкіркою (іноді – і над нижньою) міститься стовпчаста паренхіма, яка складається з клітин правильної форми (призматичної), розташованих вертикально кількома шарами і щільно прилягають одна до одної. Пухка паренхіма знаходиться під стовпчастою і над нижньою шкіркою, складається з клітин неправильної форми, які щільно не прилягають одна до одної і мають великі міжклітини, заповнені повітрям. Міжклітинники займають до 25% обсягу аркуша. Вони з'єднуються з продихами і забезпечують газообмін і транспірацію листа. Вважається, що більш інтенсивні процеси фотосинтезу відбуваються в палісадній паренхімі, оскільки її клітини мають більше хлоропластів. У клітинах пухкої паренхіми хлоропластів значно менше. У них активно запасається крохмаль та деякі інші поживні речовини.
Крізь тканини паренхіми проходять судинноволокнисті пучки (жилки). До їх складу входять провідна тканина - судини (у найдрібніших жилках - трахеїди) і ситоподібні трубки - і механічна. Зверху судинно-волокнистого пучка розташована ксилема, а знизу – флоема. По ситоподібних трубках протікають органічні речовини, що утворилися в процесі фотосинтезу, до всіх органів рослини. По судин і трахеїдів до листа надходить вода з розчиненими у ній мінеральними речовинами. Механічна тканина надає міцності листової пластинки, опору провідної тканини. Між провідною системою та мезофілом знаходиться вільний простір або апопласт .
Видозміни листа
Видозміни листя (метаморфози) виникають при виконанні додаткових функцій.
Вусики
Дозволяють рослині (горох, вика) чіплятися за предмети та закріплювати стебло у вертикальному положенні.
Колючки
Виникають у рослин, які ростуть у посушливих місцях (кактус, барбарис). У робінії псевдоакації (білої акації) колючки - це видозміни прилистків.
Лусочки
Сухі лусочки (нирок, цибулин, кореневищ) виконують захисну функцію – захищають від ушкоджень. М'ясисті лусочки (цибулини) запасають поживні речовини.
У комахоїдних рослин (росянка) листя видозмінене для уловлювання та перетравлення переважно комах.
Філодії
Це перетворення черешка в листоподібну плоску освіту.
Мінливість листа обумовлена сукупністю зовнішніх та внутрішніх факторів. Наявність в однієї рослини листя різної форми і розмірів називається гетерофілією , або різнолистістю . Спостерігається, наприклад, у водяного жовтця, стрілоліста тощо.

(Від латів. trans - крізь і spiro - дихаю). Це виведення рослиною водяної пари (випаровування води). Рослини поглинають багато води, але використовують лише її незначну частину. Воду випаровують усі частини рослини, але особливо – листя. Завдяки випаровуванню навколо рослини виникає особливий мікроклімат.
Види транспірації
Розрізняють два види транспірації: кутикулярну та устьичну.
Кутикулярна транспірація
Кутикулярна транспірація - це випаровування води всією поверхнею рослини.
Устьична транспірація
Устьична транспірація- Це випаровування води через продихи. Найбільш інтенсивною є устьична. Продихання регулюють швидкість випаровування води. Кількість продихів у різних видіврослин різне.
Транспірація сприяє надходженню нової кількості води до кореня, підняттю води по стеблі до листя (за допомогою всмоктуючої сили). Таким чином коренева системаутворює нижній водяний насос, а листя – верхній водяний насос.
Одним із факторів, що визначають швидкість випаровування, є вологість повітря: чим вона вища, тим менше випаровування (випаровування припиняється при насиченні повітря водяною парою).
Значення випаровування води: знижує температуру рослини та захищає її від перегріву, забезпечує висхідний струм речовин від кореня до надземної частини рослини. Від інтенсивності транспірації залежить інтенсивність фотосинтезів, оскільки обидва ці процеси регулюються устьичним апаратом.

Це одночасне скидання листя на період несприятливих умов. Основними причинами листопада є зміна тривалості світлового дня, зниження температури. При цьому посилюється відтік органічних речовин з листка до стебла та кореня. Спостерігається восени (іноді, посушливі роки, влітку). Листопад є пристосуванням рослини для захисту від надмірної втрати води. Разом з листям видаляються різні шкідливі продукти обміну речовин, які відкладаються (наприклад, кристали оксалату кальцію).
Підготовка до листопаду починається ще до настання несприятливого періоду. Зниження температури повітря призводить до руйнування хлорофілу. Інші пігменти стають помітними (каротини, ксантофіли), тому листя змінює забарвлення.
Клітини черешка біля стебла починають посилено ділитися і утворюють поперек його. віддільний шар із паренхіми, який легко розшаровується. Вони стають округлими, гладкими. Між ними виникають великі міжклітини, які дозволяють клітинам легко відокремлюватися. Лист залишається прикріпленим до стебла лише завдяки судинно-волокнистим пучкам. На поверхні майбутнього листового рубця заздалегідь утворюється захисний шар пробкової тканини.
У однодольних рослин та трав'янистих дводольних не утворюється відокремлювальний шар. Лист відмирає, поступово руйнується, залишаючись на стеблі.
Опале листя розкладається ґрунтовими мікроорганізмами, грибами, тваринами.


